
Motor and Somatosensory Conversion Disorder: A Functional Unawareness Syndrome?
Abstract
Although conversion disorder is closely connected to the origins of neurology and psychiatry, it remains poorly understood. In this article, the authors discuss neural and clinical parallels between lesional unawareness disorders and unilateral motor and somatosensory conversion disorder, emphasizing functional neuroimaging/disease correlates. Authors suggest that a functional-unawareness neurobiological framework, mediated by right hemisphere-lateralized, large-scale brain network dysfunction, may play a significant role in the neurobiology of conversion disorder. The perigenual anterior cingulate and the posterior parietal cortices are detailed as important in disease pathophysiology. Further investigations will refine the functional-unawareness concept, clarify the role of affective circuits, and delineate the process through which functional neurologic symptoms emerge.
Conversion disorder is a neuropsychiatric condition at the interface of neurology and psychiatry. Clinically trained neurologists see approximately 30% of outpatients for medically unexplained illness,1 and up to 18% of patients with unexplained illness are diagnosed with functional neurologic symptoms.2 Despite frequent clinical exposure to conversion disorder, after identifying functional signs, including distractibility and inconsistency, without objective deficits, neurologists are uncomfortable in the psychiatrist’s chair.3 The lack of a conceptual framework through which to understand this disorder is exemplified by a neurologist’s stating “Well, I don’t really know….I suppose it may be their way of dealing with problems they can’t solve”3. It is the collective clinical experience of the authors that part of the challenge for physicians in working with these patients lies in the absence of an accepted neurobiological framework through which to understand the clinical phenotype of functional neurological symptoms.
Advanced functional neuroimaging techniques now allow access to neural-system dysfunction in conversion disorder. Despite multiple neuroimaging studies and reviews on the topic,4 a brain-based conceptual model through which to understand functional neurological disturbances has yet to be recognized. In this article, conversion disorder is first historically contextualized to allow for the integration of neurologic and psychiatric concepts with an emerging neurobiology. Second, the neurobiology of the disorder is explored by reviewing functional neuroimaging findings in the most well-studied subset of patients (functional unilateral motor and somatosensory disturbances). Thereafter, conversion disorder is positioned among the “unawareness” disorders in neuropsychiatry, and functional unawareness is suggested as a useful neurobiological framework through which to understand this illness.
In this article, the newly proposed Diagnostic and Statistical Manual of Mental Disorders (DSM-5) nomenclature is adopted, and conversion disorder is referred to as functional neurological disorder (FND).5
Approach
The purpose of this article is to offer a neurobiological perspective through which clinicians and translational scientists may begin to understand patients with functional unilateral motor and somatosensory disturbances, rather than provide a comprehensive review of published neuroimaging studies on this enigmatic disease. To this end, we searched PubMed and Google Scholar for relevant articles in “conversion disorder” and “hysteria” published between January 1995 and December 2010; emphasis was placed on articles using Technetium-99 single photon-emission computed tomography (99Tc-SPECT), fluorodeoxy-glucose positron emission tomography (FDG-PET), and functional magnetic resonance imaging (fMRI) techniques. Analyses exploring hypnosis or feigning behaviors in comparison to FND were omitted, to limit comparisons between FND and additional incompletely-understood biological phenomena.
Historical Perspective: Origins of Hysteria
FND was first known in the medical literature as “hysteria,” and it has engaged prominent individuals in the history of medicine, neurology, and psychiatry. Although initially described in ancient Greece with gynecologic (“wandering womb”) themes and later as demonic possession after the rise of Christian civilization,6 hysteria was medicalized in the 19th century by the French neurologist Jean-Martin Charcot, through his study of patients at the Salpêtrière Hospital. Charcot recognized hysteria as an acceptable, neurologically diagnosable condition, stating “the neurological tree has its branches; neurasthenia, hysteria, epilepsy, all the types of mental conditions, progressive paralysis, gait ataxia.”7
Reclassification of hysteria as a psychiatric disorder began with the writings of Sigmund Freud, an Austrian neurologist and founder of psychoanalysis. Freud argued, “the aetiology was to be sought in sexual factors.”8 He coined the term “conversion hysteria” and described a process whereby “the affective idea is converted into a physical phenomenon.”8 Whereas Freud described a transformation of psychic conflict into somatic symptoms, Pierre Janet, a French psychologist, suggested that “hysteria is a form of mental depression characterized by retraction of the field of personal consciousness and a tendency to the dissociation and emancipation of the system of ideas and functions that constitute personality.”9 Thus, by the late 19th/early 20th century, FND was an accepted neurologic and psychiatric condition.
The FND clinical syndrome is now recognized to occur more often in women, with symptom onset often presenting during the teens or early-20s. Patients have an elevated rate of Axis I comorbidity, and symptoms are generally acute in onset, of short duration with multiple reoccurrences, and commonly occur after a psychological stress. As originally described by Freud, patients may exhibit an unusually calm demeanor regarding their symptoms (“la belle indifférence”), whereas others exhibit a high degree of emotionality. Childhood sexual and physical trauma may be reported. In functional weakness, movements, if performed, are slow, tentative, and nonsustained. Functional somatosensory deficits may manifest with nonphysiological demarcations, such as immediately left of the sternum, or may exhibit fluctuating boundaries.
Neural formulations of FND have emerged in the last few decades. Whitlock, in 1967, described hysteria as a neuropsychiatric disorder involving attentional dysregulation, characterized by “selective depression of awareness of a bodily function.”10 Sierra and Berrios, in 1999, proposed altered attention and awareness, mediated in part by inferior parietal cortex dysfunction, as a neurobiological model for FND.11 Lateralized right-hemispheric dysfunction with relative preservation of a narrative, interpretative left hemisphere, has also been suggested as a neural explanation for FND, and implies an overlap with delusional disorders.12,13 Also, theories applied to the broader category of somatoform disorders have emphasized affect dysregulation and somatic amplification as important mediators of disease.14,15
Neuroimaging of Unilateral Motor/Somatosensory FND
Over the past 10–15 years, the neurobiology of unilateral motor and somatosensory functional neurologic disturbances has advanced with the use of functional neuroimaging techniques, including 99Tc-SPECT, FDG-PET, and fMRI. Prefrontal inhibition of primary motor/somatosensory cortex,16–19 intentional disturbances,20,21 attentional dysregulation,22–24 impaired action authorship recognition,25 and affective disturbances26,27 are major neural processes implicated in the neuroscience of FND.
Inhibition
The first published neuroimaging study in FND was performed on a 32-year-old woman with panic attacks, depression, and recent marital discord, who developed left-sided weakness (preserved finger movements, with arm weakness and foot clumsiness) and paresthesias.1899Tc-SPECT blood flow patterns in response to left median nerve stimulation revealed right frontal cortex hyperperfusion and right parietal cortex (including primary somatosensory cortex) hypoperfusion, only during symptomatic periods. In related studies, medial prefrontal cortex (PFC) hyperactivation was replicated and further localized to the anterior cingulate cortex (ACC), orbitofrontal cortex (OFC), and ventromedial PFC.16,17,19 Medial PFC inhibition of unimodal primary somatosensory and motor cortices was postulated to explain functional numbness and weakness.17,18
Intention
Impairment in the generation of motor intention was also hypothesized for patients with FND.20 Three men with functional arm weakness (two, left-sided) and previous depression exhibited left dorsolateral PFC hypometabolism compared with healthy subjects on FDG-PET during performance of joy stick movements. Functional weakness was suggested to reflect a disturbance in motor intention planning. This notion has been supported by evidence of altered functional connectivity between the dorsolateral PFC and sensorimotor areas in patients with FND,21 and further delineation of a role for the dorsolateral PFC in motor intention generation and cognitive control of motor behavior.28,29
Attention
A third neurocognitive function, attention, was explored, using 99Tc-SPECT in seven patients with functional unilateral hemiparesis/hypoesthesia (4, left-sided; 5, with depressed mood) during rest and passive bilateral vibratory stimulation.24 Contralateral thalamic and basal-ganglia blood-flow reductions were observed only in symptomatic patients during sensory stimulation. Impairments in striato-thalamic components of attentional and motivational neural networks were proposed as mediators of FND. Also, in related investigations,30 patients with unilateral motor FND exhibited bilateral striatal/pallidal and right-thalamic volume reductions. Thalamic dysfunction was replicated in functional anesthesia,23 and a lack of striato-thalamic circuit activation was also found in patients with functional weakness.19
In a unique case, a 56-year-old, right-handed woman with emotional distress related to marital discord presented with transient functional left arm and leg weakness, hypoesthesia and functional left-hemispatial neglect.31 fMRI obtained while she was symptomatic, during performance of a line-bisection task, showed right ACC hyperactivity relative to healthy subjects. Thus the perigenual ACC (pACC), posterior parietal cortex (PPC),18 striatum, and thalamus, regions implicated in attentional neurobiology, exhibited dysfunctional activity in FND patients.
Action Authorship
Disturbances of motor intention awareness and self-agency have also been suggested for patients with FND. Voon and colleagues25 examined fMRI blood-oxygen level-dependent (BOLD) patterns in eight FND patients (two with major depression, three with generalized anxiety disorder) with positional, predominantly unilateral, unexplained tremors as compared with volitional movements. The right temporoparietal junction (TPJ) was less active during unexplained movements, and reduced functional connectivity occurred between the right TPJ and bilateral sensory/motor cortex, ACC, medial PFC, and right superior parietal lobule. Given the roles of the TPJ and adjacent regions (PPC/superior temporal gyrus) in motor intention awareness and self-agency perception,32–35 diminished right TPJ activity and aberrant connectivity were suggested to explain FND patients’ inability to recognize themselves as the authors of their actions.
Affective Disturbances
Two imaging studies directly probed affective (limbic) circuit dysfunction in FND patients. A scripted, traumatic memory provocation fMRI study was administered to a woman with selective amnesia for a romantic break-up and functional right hemiparesis/hypoesthesia. fMRI showed right amygdala, ACC, parietal cortex, and inferior frontal activations during cued recall of repressed memories, as compared with readily-recalled events.26 Amygdalar activation was proposed as a biomarker of heightened emotional salience for repressed memories. A second fMRI study examined affective facial processing in 16 mixed-phenotype, hyperkinetic, FND patients with anxiety and depression, compared with healthy subjects. Patients with hyperkinetic FND showed increased amygdalar activation to happy faces and increased amygdalar functional connectivity with the supplementary motor area (SMA).27 Motor disinhibition resulting from heightened limbic–SMA interactions via striato-pallidal-thalamic projections were proposed as mechanisms of limbic influences on motor actions.25 These studies supported the idea of amygdalar and ACC dysfunction in patients with FND.
Having reviewed prefrontal-mediated inhibition, intentional disturbances, inattention, impaired action-authorship recognition, and affective dysregulation as major neural processes implicated in the neurobiology of functional neurologic disturbances, the following section frames unilateral functional motor and somatosensory disturbances as “disorders of unawareness.”
Unawareness and Right-Hemisphere Dysfunction
The neglect syndrome is defined as a right-brain/left-body unawareness disorder characterized by impaired ability to report, respond, or orient to novel or salient stimuli; it is associated with right-hemispheric lesions.36,37 Subtypes include sensory (somatosensory, visual, auditory), motor, hemispatial, and personal neglect.38 Of particular interest is motor neglect, characterized by impaired motor intention generation and exemplified by limb underuse, hypokinesia, and inability to sustain motor movements (motor impersistence), despite the absence of corticospinal system damage.38,39 Notably, functional weakness resembles motor neglect. Structural studies in motor-neglect stroke patients identify a pattern of right hemisphere-predominant frontal, parietal, striatal and thalamic lesions,40–44 and implicate cortico-cortical (frontal-parietal) and cortico-subcortical pathways in lesional motor neglect.
Concepts related to motor neglect include motor intention awareness (the conscious recognition of the desire to move) and self-agency (the sense that we are the authors of our own movements).34 Early behavioral studies examining predictions of motor action45 and hand position46 suggested that individuals used internal sensory predictions to anticipate motor actions, a process termed forward modeling or corollary discharge.32,45–47 Forward modeling allows the central nervous system to maintain accurate performance based on the predicted sensory consequences before the actual processing of sensory afferent signals. Furthermore, observations have shown that the predictability of sensory consequences following motor actions correlates with perceived self-agency.48 Forward modeling, awareness of motor intention, and self-agency may be related concepts that involve common or interrelated neural circuits. Also, aberrant forward modeling has been extended to other modalities and proposed as an explanation for neuropsychiatric disturbances, including auditory hallucinations and delusions of control in schizophrenic patients experiencing their own internal dialogue or actions as “other.”47
A role for the PPC in forward modeling has been suggested by lesional and functional neuroimaging studies. Lesions of the superior parietal lobule have been linked to time-dependent decrements in sustained arm motor function and proprioceptive unawareness,49 and updates of limb posture recruited bilateral superior parietal cortex activations in an fMRI spatial-pointing task.50 These results supported a role for the PPC, particularly the superior parietal lobule, in real-time dynamic internal sensorimotor integration, a prerequisite for predictive forward-modeling.
Studies also implicated the PPC in motor intention awareness and self-agency. Patients with right inferior parietal lobule (IPL) lesions exhibited a shortened latency between conscious recognition of impending action and motor execution. This suggested a deficit in motor intention awareness before the imminent release of motor action.34,35 Interestingly, patients undergoing intraoperative IPL cortical stimulation experienced the conscious desire for motor action at low intensity and an experience of motor accomplishment at high intensity.33 This implied that perception of action authorship involved similar neural networks to that of motor intention awareness. In further support of this concept, patients with IPL lesions falsely attributed examiner-made, complex hand movements as their own,51 and bilateral angular gyrus activations occurred in individuals making agency determinations.52 Thus, the PPC, a node in the large-scale brain network disturbed in lesional neglect (including motor neglect), participates in forward-modeling, motor-intention awareness, and self-agency perceptions.34,53,54
Somatosensory neglect (e.g. tactile extinction to double simultaneous stimulation in the absence of lateralized somatosensory deficits) is difficult to isolate clinically, and frequently coexists with primary somatosensory disturbances, motor weakness, and hemispatial neglect, limiting its study in isolation. However, extrapolation from hemispatial neglect, the most studied neglect syndrome subtype, allows an inference into the neurobiology of somatosensory unawareness. Primarily on the basis of lesional analyses, a distributed, right hemisphere dominant, cortical and subcortical network for hemispatial neglect has been suggested to include the PPC, TPJ, superior temporal gyrus, ACC, frontal eye fields, inferior frontal gyrus, striatum, and thalamus.55–63 Somatosensory and motor neglect, therefore, may share similar neural substrates.
FND: A Functional Unawareness Syndrome
Functional neural circuit disturbances in motor and somatosensory FND overlap with the cortico-cortical and cortico-subcortical pathways implicated in lesional motor and somatosensory neglect. A number of cross paradigm studies in patients with functional limb weakness demonstrated patterns of neural dysfunction in right-greater-than-left PPC (extended to include the TPJ),18,25 ACC,17,26,64 striatum, and thalamus.19 Functional neuroimaging studies in patients with functional somatosensory deficits exhibited similar dysfunction in attentional regions, again including right-greater-than-left parietal cortex,18 ACC,17,23,26,31 striatum,24 and thalamus.23,24 In further support of an overlapping neural substrate between FND and lesional motor and somatosensory unawareness, a case of transient functional left-of-midline hemibody anesthesia was described in a patient with a right parietal infarct.65 Psychogenic non-epileptic seizures may occur more often in patients with right-hemispheric lesions or right-hemispheric dysfunction on electroencephalogram.66 Lastly, left-sided bodily deficits have traditionally been considered more frequent in FND,67 although a recent metaanalysis failed to show this effect.68
Using a best-fit approach to synthesize neuroimaging and phenomenological data, functional somatosensory unawareness and functional motor unawareness are suggested as major contributors in the pathophysiology of unilateral somatosensory and motor related FND (Figure 1). Clinical and neural features of functional disturbances share a common denominator with lesional neglect: unawareness. Individual variation across unilateral motor and somatosensory FND phenotypes may be accounted for, in part, by relative contributions of prefrontal, posterior parietal, and subcortical components. Furthermore, pACC-subcortical and PPC-subcortical pathways may provide complementary, but distinct, unawareness contributions.69,70 Disturbances of motivated behavior/motor control/affect regulation may arise preferentially from pACC–subcortical pathways (including pACC–amygdalar circuits), whereas attentional and perceptual miscalculations may arise from PPC–subcortical dysfunction.69,71–73 Reciprocal cortico–cortical connections among the pACC, PPC, and dorsolateral PFC facilitate interactions among circuits mediating affect regulation, awareness, intention, and cognitive control.74,75
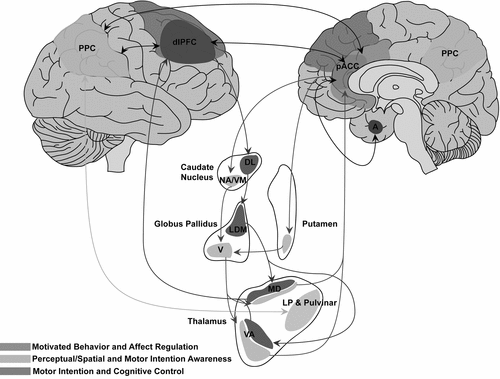
Dysfunction in the perigenual anterior cingulate cortex (pACC) and its subcortical connections (including reciprocal cingulate–amygdalar connections) results preferentially in impaired motivated behavior, motor control, and/or affect regulation. Dysfunction in posterior parietal cortex (PCC) and its subcortical connections results preferentially in impaired spatial and perceptual awareness, including aberrant forward modeling, motor intention awareness, and/or self-agency. Reciprocal cortico–cortical connections among the pACC, PCC, and the dorsolateral prefrontal cortex (dlPFC) facilitate interactions between awareness and intentional, cognitive control circuits.
VM: ventromedial; DL: dorsolateral; NA: nucleus accumbens; VA: ventral anterior; LP: lateral posterior; MD: mediodorsal; LDM: lateral dorsomedial; V: ventral; A: amygdala.
In this conceptualization, in addition to the PPC, an important role is postulated for the pACC in functional motor/somatosensory neglect, given its dual cognitive and emotional functions mediated by structural connections with the posterior parietal, premotor, dorsolateral prefrontal, OFC, subgenual ACC (sgACC), and medial-temporal cortices.76–80 Human and animal models also implicate the ACC in stress-related, maladaptive experience-dependent neuroplastic change. For example, patients with posttraumatic stress disorder (PTSD) after childhood sexual trauma exhibited functional and morphologic abnormalities in the ACC, as compared with healthy subjects.81,82 Chronic stress in animal models induced dendritic spine reduction in the ACC and hippocampus, with parallel enhanced dendritic arborization in the amygdala.83,84 Reciprocal top-down (ACC) and bottom-up (amygdala) sites of aberrant experience-dependent neuroplastic change, and medial/lateral and dorsal/ventral PFC interactions specific to FND patients require more exploration in the context of extended emotional/affective regulation–neural circuit explorations (Figure 2). Furthermore, the OFC, implicated in social-emotional evaluation and behavioral control in the context of changing contingencies,85 and the sgACC, implicated in the modulation of automatic emotional behavior,86 are additional paralimbic regions that require further investigation. Initial findings, nonetheless, position the pACC and related networks as mediators of impaired affect-regulation and cognitive processes in patients with FND.
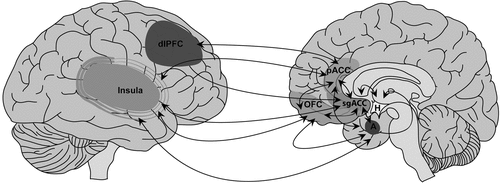
Reciprocal connections are outlined among the perigenual anterior cingulate cortex (pACC), subgenual ACC (sgACC), orbitofrontal cortex (OFC), dorsolateral prefrontal cortex (dlPFC), insula, amygdala (A), and hypothalamus (H). Parallel ACC, dlPFC and OFC prefrontal-subcortical pathways (not shown) also require more exploration in the context of studies probing affective regulation in patients with functional neurological disorder (FND).
Several important implications arise from the proposed neurobiological framework. The outlined cortico-cortical and cortical-subcortical pathways allow clinicians to understand the presentation of functional symptoms independent from notions of psychic tension and the unconscious mind; functional deficits in-and-of-themselves may not necessarily be symbolic of the particular emotional stress experienced by the patient, but rather may be the product of intrinsic neural connectivity patterns. Thus, the basic form or phenotype in which functional symptoms present may be driven by intrinsic neural connections, whereas the specific context of functional symptoms (i.e., why and when symptoms occur) may be a product of interactions between the patients’ psychosocial stressors and attentional, limbic/paralimbic, and sensory-motor cortices.
Also, neurologists are frequently frustrated by interactions with FND patients, since clinicians may view them as fabricating their symptoms. This frustration is markedly less when treating unawareness in right-hemisphere stroke patients with neglect. Conceptualizing unilateral functional motor and somatosensory deficits as functional neglect or unawareness, rather than willful deception or simulation, may improve the therapeutic alliance between physician and patient, and potentially increase the number of clinicians inclined to work longitudinally with FND patients. Importantly, this framework does not, in our opinion, undervalue the role of affective disturbances in the presentation of functional neurologic symptoms, but rather contextualizes the patients’ presentation in terms readily understood by clinicians. Finally, the identification of cortical sites, including the PPC and the dorsolateral PFC, offers targets to investigate for their therapeutic intervention in patients with FND; these include the potential use of neuromodulation techniques (i.e., transcranial magnetic stimulation) to modify aberrant functional activation patterns.
Fixed, False Illness Beliefs in FND: A Left-Hemisphere Delusion
Having suggested functional unawareness as a neurobiological framework through which to understand motor and somatosensory FND, inherent differences in right-brain/left-brain functionality may clarify the fixed beliefs held by FND patients regarding the nature of their illness. Investigations in split-brain patients identified a role for the left hemisphere in narrative, interpreter-like functions.87 Together with evidence that right-hemisphere lesions predisposed to delusions,88 right-hemisphere dysfunction with relative preservation of left lateralized function has been proposed as a neural basis of fixed, false beliefs, including delusional misidentification syndromes.12,13 Unawareness driven predominantly by right-hemisphere dysfunction with preserved interpreter functions in the left hemisphere may result in a functional interhemispheric disconnection, and it helps explain the persistent, false beliefs held by FND patients regarding their disease.
FND and Dissociation: Emotional Unawareness?
Using the functional unawareness concept, it is intriguing to emphasize emotional unawareness a century after Pierre Janet offered restriction of the field of consciousness (dissociation) as a theory through which to understand FND.9 It has been suggested that the right hemisphere plays a dominant role in emotional expression, a concept supported by evidence linking right-hemisphere lesions with flat affect, aprosody, and blunted autonomic responses.89–91 Dissociative tendencies are also now well recognized in FND by use of standardized self-report scales such as the Dissociative Experience Scale.64 Although specific neuroimaging studies correlating dissociative tendencies with functional activation patterns in patients with FND are lacking, studies in related conditions with high rates of dissociation, including depersonalization disorder92 and PTSD,93 suggest a role for the right PPC, particularly the precuneus, and the ACC in dissociative neurobiology. Direct exploration of dissociation and alexithymia in FND populations may help clarify mechanisms of emotional unawareness and impaired emotional expression.
Limitations and Other Considerations
There are several important limitations to address regarding the suggested conceptual framework. The discussion is limited to unilateral functional motor and somatosensory disturbances and does not incorporate other FND subtypes, including astasia-abasia (functional gait disorder),94 functional blindness,95 and psychogenic non-epileptic seizures. All FND subtypes may not have the same mechanistic network explanations for distinct phenotypes, and research in these and other subtypes warrant further investigation. Also, many of the functional neuroimaging studies cited examine single cases18,26,31 and case–control studies with a small number of subjects,20,24,25 increasing the possibility of type I statistical errors. Negative affective disturbances were also not consistently controlled for methodologically across studies, suggesting potential confounds for the prefrontal, subcortical, and limbic findings.
Comparisons with deliberate feigning or hypnosis-induced functional disturbances are also omitted. Although deliberate feigning has been used as a comparison condition in some FND neuroimaging studies,19,20,96 the neural correlates of feigning are not yet well understood, which adds increased difficulty when comparing the neural activation patterns to patients with FND. Hypnosis-related research is also not incorporated in the FND framework, since hypnosis requires more exploration, but we acknowledge the possibility of an overlap in the biology of hypnosis and FND.97
The functional-unawareness concept may also be interpreted with evolving psychological models. It is important, however, first to note that, when considering the integration of psychological and neural models, researchers should not necessarily search for the neural correlates of a given psychological concept, which may or may not be supported by empirical evidence. With this in consideration, one possible psychodynamic formulation invokes mechanisms of adaptive and maladaptive psychological defense.98 Defense mechanisms are conceptualized as the ego’s efforts to cope with psychological stress, and pathologic somatic-based defenses thought to be used by patients with FND would reduce transpsychic conflict through bodily displacement. The ACC has been implicated in emotional expression/regulation, maladaptive neuroplasticity, and conflict monitoring (cognitive neuroscience definition), and offers intriguing possibilities for integrative synthesis. However, more research is needed to fully integrate psychological and neurobiological concepts in patients with FND.
Conceptualization of FND as a disorder of unawareness also does not explain differences between hypokinetic and hyperkinetic functional motor symptoms. Inhibitory or disinhibitory effects of premotor regions (i.e., SMA) on primary motor cortices, and modulated by interactions with the PPC, dorsolateral PFC, and limbic/paralimbic regions, may help clarify phenotypic differences.27,34 This may be analogous to differences in alien hand syndrome phenotypes; medial-frontal lesions result in hypermotor, foreign behaviors, whereas posterior variants present more commonly with disturbances of posture and levitation.99 Also, further clarification is needed of the role of the left hemisphere in unawareness, as seen infrequently, for example, in left-hemisphere lesions resulting in contralesional visual-spatial neglect or unawareness of deficit in Wernicke’s aphasia.36,37 Lastly, this article reconciles many of the distinct and overlapping neural-circuit findings across paradigms in FND, but falls short of providing a unifying neurobiological model incorporating the entire body of unilateral motor and somatosensory FND literature.96,100 More detailed understanding will require larger subject groups, multiparadigm neuroimaging studies (including neuroimaging at rest to explore default-mode network disturbances), and non-imaging, cross-disciplinary research to provide a fully integrative model.
Conclusions
In summary, FND has served an important role in the origins of both neurology and psychiatry. In this article, we identify overlapping neural circuitry between lesional motor/somatosensory neglect and some forms of FND. Functional motor unawareness and functional somatosensory unawareness are suggested as conceptual frameworks through which to understand functional motor and somatosensory neurologic symptoms. Early postulations by Charcot delineating FND as a functional neurologic disorder and Janet’s dissociation theory for unawareness in hysteria are supported by evolving FND neural circuit delineations. Increasingly well-characterized large-scale brain networks mediating cognitive and affective processes and modulated by experience-dependent neuroplasticity provide the foundation for a neurobiological conceptualization through which to understand functional neurologic disturbances. Further investigation is needed to test and refine the functional-unawareness concept, delineate the biological processes through which functional symptoms arise, and clarify the role of affective neural circuits in FND biology. Subsequent investigation and synthesis will enable the development of a unified model for functional neurologic disturbances.
1 : The outcome of neurology outpatients with medically unexplained symptoms: a prospective cohort study. J Neurol Neurosurg Psychiatry 2003; 74:897–900Crossref, Medline, Google Scholar
2 : Symptoms ‘unexplained by organic disease’ in 1144 new neurology out-patients: how often does the diagnosis change at follow-up? Brain 2009; 132:2878–2888Crossref, Medline, Google Scholar
3 : In the psychiatrist’s chair: how neurologists understand conversion disorder. Brain 2009; 132:2889–2896Crossref, Medline, Google Scholar
4 : Dissociation in hysteria and hypnosis: evidence from cognitive neuroscience. J Neurol Neurosurg Psychiatry 2010; 82:332–339Crossref, Medline, Google Scholar
5 : Issues for DSM-5: Conversion disorder. Am J Psychiatry 2010; 167:626–627Crossref, Medline, Google Scholar
6 : Freud and the great neurosis: discussion paper. J R Soc Med 1983; 76:57–61Crossref, Medline, Google Scholar
7 : Leçons du mardi á la Salpêtrière: policliniques, 1887–1888. Paris, Bureaux du Prográes Mâedical, 1887Google Scholar
8 : Studies on hysteria. London, Hogarth Press, 1956Google Scholar
9 : The major symptoms of hysteria; fifteen lectures given in the Medical School of Harvard University. New York, Macmillan, 1907Crossref, Google Scholar
10 : The aetiology of hysteria. Acta Psychiatr Scand 1967; 43:144–162Crossref, Medline, Google Scholar
11 : Towards a Neuropsychiatry of Conversive Hysteria. Cogn Neuropsychiatry 1999; 4:267–287Crossref, Google Scholar
12 : Right Cerebral Hemisphere Dominance for a Sense of Corporeal and Emotional Self. Epilepsy Behav 2000; 1:60–73Crossref, Google Scholar
13 : Delusional misidentifications and duplications: right brain lesions, left brain delusions. Neurology 2009; 72:80–87Crossref, Medline, Google Scholar
14 : Functional somatic syndromes. Ann Intern Med 1999; 130:910–921Crossref, Medline, Google Scholar
15 : Somatoform disorders as disorders of affect regulation: a development perspective. Int Rev Psychiatry 2006; 18:13–24Crossref, Medline, Google Scholar
16 : Increased self-monitoring during imagined movements in conversion paralysis. Neuropsychologia 2007; 45:2051–2058Crossref, Medline, Google Scholar
17 : The functional anatomy of a hysterical paralysis. Cognition 1997; 64:B1–B8Crossref, Medline, Google Scholar
18 : Altered cerebral blood flow during hysterical paresthesia. Biol Psychiatry 1995; 37:134–135Crossref, Medline, Google Scholar
19 : Motor inhibition in hysterical conversion paralysis. Neuroimage 2009; 47:1026–1037Crossref, Medline, Google Scholar
20 : Discrete neurophysiological correlates in prefrontal cortex during hysterical and feigned disorder of movement. Lancet 2000; 355:1243–1244Crossref, Medline, Google Scholar
21 : Altered connectivity between prefrontal and sensorimotor cortex in conversion paralysis. Neuropsychologia 2010; 48:1782–1788Crossref, Medline, Google Scholar
22 : Unexplained neurologic symptoms: an fMRI study of sensory conversion disorder. Neurology 2006; 67:2036–2038Crossref, Medline, Google Scholar
23 : Altered central somatosensory processing in chronic pain patients with “hysterical” anesthesia. Neurology 2003; 60:1501–1507Crossref, Medline, Google Scholar
24 : Functional neuroanatomical correlates of hysterical sensorimotor loss. Brain 2001; 124:1077–1090Crossref, Medline, Google Scholar
25 : The involuntary nature of conversion disorder. Neurology 2010; 74:223–228Crossref, Medline, Google Scholar
26 : Imaging repressed memories in motor conversion disorder. Psychosom Med 2007; 69:202–205Crossref, Medline, Google Scholar
27 : Emotional stimuli and motor conversion disorder. Brain 2010; 133:1526–1536Crossref, Medline, Google Scholar
28 : Unconscious determinants of free decisions in the human brain. Nat Neurosci 2008; 11:543–545Crossref, Medline, Google Scholar
29 : Functional anatomy of motor urgency. Neuroimage 2007; 37:243–252Crossref, Medline, Google Scholar
30 : Volumetric investigation of brain regions in patients with conversion disorder. Prog Neuropsychopharmacol Biol Psychiatry 2006; 30:708–713Crossref, Medline, Google Scholar
31 : Functional brain imaging in a woman with spatial neglect due to conversion disorder. JAMA 2009; 302:2552–2554Crossref, Medline, Google Scholar
32 : Action prediction in the cerebellum and in the parietal lobe. Exp Brain Res 2003; 153:239–245Crossref, Medline, Google Scholar
33 : Movement intention after parietal cortex stimulation in humans. Science 2009; 324:811–813Crossref, Medline, Google Scholar
34 : A parietal-premotor network for movement intention and motor awareness. Trends Cogn Sci 2009; 13:411–419Crossref, Medline, Google Scholar
35 : Altered awareness of voluntary action after damage to the parietal cortex. Nat Neurosci 2003; 7:80–84Crossref, Medline, Google Scholar
36 : Right hemisphere dominance for attention: the mechanism underlying hemispheric asymmetries of inattention (neglect). Neurology 1980; 30:327–330Crossref, Medline, Google Scholar
37 : A cortical network for directed attention and unilateral neglect. Ann Neurol 1981; 10:309–325Crossref, Medline, Google Scholar
38 : Neglect and related disorders. Semin Neurol 2000; 20:463–470Crossref, Medline, Google Scholar
39 : Anosognosia for hemiplegia: an electrophysiologic investigation of the feed-forward hypothesis. Neurology 1994; 44:1804–1808Crossref, Medline, Google Scholar
40 : Motor neglect associated with loss of action inhibition. J Neurol Neurosurg Psychiatry 2008; 79:1401–1404Crossref, Medline, Google Scholar
41 : Right motor neglect associated with dynamic aphasia, loss of drive and amnesia: case report and cerebral blood flow study. Neuropsychologia 1992; 30:109–121Crossref, Medline, Google Scholar
42 : Motor neglect. J Neurol Neurosurg Psychiatry 1983; 46:152–158Crossref, Medline, Google Scholar
43 : Motor neglect associated with a discrete parietal lesion. Neurology 1994; 44:1164–1166Crossref, Medline, Google Scholar
44 : Unilateral hypokinesia and motor extinction. Neurology 1981; 31:445–448Crossref, Medline, Google Scholar
45 : Predicting the consequences of our own actions: the role of sensorimotor context estimation. J Neurosci 1998; 18:7511–7518Crossref, Medline, Google Scholar
46 : An internal model for sensorimotor integration. Science 1995; 269:1880–1882Crossref, Medline, Google Scholar
47 : Neuropsychology of schizophrenia, what are the implications of intellectual and experiential abnormalities for the neurobiology of schizophrenia? Br Med Bull 1996; 52:618–626Crossref, Medline, Google Scholar
48 : Illusion of sense of self-agency: discrepancy between the predicted and actual sensory consequences of actions modulates the sense of self-agency, but not the sense of self-ownership. Cognition 2005; 94:241–255Crossref, Medline, Google Scholar
49 : Maintaining internal representations: the role of the human superior parietal lobe. Nat Neurosci 1998; 1:529–533Crossref, Medline, Google Scholar
50 : Parietal updating of limb posture: an event-related fMRI study. Neuropsychologia 2006; 44:2685–2690Crossref, Medline, Google Scholar
51 : Perception of self-generated movement following left parietal lesion. Brain 1999; 122:1867–1874Crossref, Medline, Google Scholar
52 : The angular gyrus computes action awareness representations. Cereb Cortex 2007; 18:254–261Crossref, Medline, Google Scholar
53 : Control of goal-directed and stimulus-driven attention in the brain. Nat Rev Neurosci 2002; 3:201–215Crossref, Medline, Google Scholar
54 : Spatial attention and neglect: parietal, frontal and cingulate contributions to the mental representation and attentional targeting of salient extrapersonal events. Philos Trans R Soc Lond B Biol Sci 1999; 354:1325–1346Crossref, Medline, Google Scholar
55 : Brain–behavior correlations in hemispatial neglect using CT and SPECT: the Sunnybrook Stroke Study. Neurology 1998; 50:901–908Crossref, Medline, Google Scholar
56 : The anatomy of visual neglect. Brain 2003; 126:1986–1997Crossref, Medline, Google Scholar
57 : The anatomy of unilateral neglect after right-hemisphere stroke lesions. A clinical/CT-scan correlation study in man. Neuropsychologia 1986; 24:609–622Crossref, Medline, Google Scholar
58 : Neglect following damage to frontal lobe or basal ganglia. Neuropsychologia 1980; 18:123–132Crossref, Medline, Google Scholar
59 : Cortical and subcortical anatomy of chronic spatial neglect following vascular damage. Behav Brain Funct 2008; 4:43–52Crossref, Medline, Google Scholar
60 : Spatial awareness is a function of the temporal not the posterior parietal lobe. Nature 2001; 411:950–953Crossref, Medline, Google Scholar
61 : The anatomy of spatial neglect based on voxelwise statistical analysis: a study of 140 patients. Cereb Cortex 2004; 14:1164–1172Crossref, Medline, Google Scholar
62 : The subcortical anatomy of human spatial neglect: putamen, caudate nucleus and pulvinar. Brain 2002; 125:350–360Crossref, Medline, Google Scholar
63 : Left unilateral neglect as a disconnection syndrome. Cereb Cortex 2007; 17:2479–2490Crossref, Medline, Google Scholar
64 : Childhood emotional abuse and dissociation in patients with conversion symptoms. Psychiatry Clin Neurosci 2009; 63:670–677Crossref, Medline, Google Scholar
65 : Conversion sensory symptoms associated with parietal lobe infarct: case report, diagnostic issues and brain mechanisms. J Psychiatry Neurosci 2002; 27:118–122Medline, Google Scholar
66 : Nondominant hemisphere lesions and conversion nonepileptic seizures. J Neuropsychiatry Clin Neurosci 2001; 13:367–373Link, Google Scholar
67 : Handedness and the lateral distribution of conversion reactions. J Nerv Ment Dis 1977; 164:122–128Crossref, Medline, Google Scholar
68 : Are functional motor and sensory symptoms really more frequent on the left? A systematic review. J Neurol Neurosurg Psychiatry 2002; 73:578–581Crossref, Medline, Google Scholar
69 : Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci 1986; 9:357–381Crossref, Medline, Google Scholar
70 : Frontal-subcortical circuits and human behavior. Arch Neurol 1993; 50:873–880Crossref, Medline, Google Scholar
71 : Non-invasive mapping of connections between human thalamus and cortex using diffusion imaging. Nat Neurosci 2003; 6:750–757Crossref, Medline, Google Scholar
72 : Striatal connections of the parietal association cortices in rhesus monkeys. J Comp Neurol 1993; 332:175–197Crossref, Medline, Google Scholar
73 : Topographic projections on the globus pallidus and the substantia nigra of selectively placed lesions in the precommissural caudate nucleus and putamen in the monkey. Exp Neurol 1971; 33:584–596Crossref, Medline, Google Scholar
74 : Posterior parietal cortex in rhesus monkey, I: parcellation of areas based on distinctive limbic and sensory corticocortical connections. J Comp Neurol 1989; 287:393–421Crossref, Medline, Google Scholar
75 : Limbic and sensory connections of the inferior parietal lobule (area PG) in the rhesus monkey: a study with a new method for horseradish peroxidase histochemistry. Brain Res 1977; 136:393–414Crossref, Medline, Google Scholar
76 : Contributions of anterior cingulate cortex to behaviour. Brain 1995; 118:279–306Crossref, Medline, Google Scholar
77 : Efferent connections of the cingulate gyrus in the rhesus monkey. Exp Brain Res 1981; 42:319–330Crossref, Medline, Google Scholar
78 : Pain and emotion interactions in subregions of the cingulate gyrus. Nat Rev Neurosci 2005; 6:533–544Crossref, Medline, Google Scholar
79 : Primate anterior cingulate cortex: where motor control, drive and cognition interface. Nat Rev Neurosci 2001; 2:417–424Crossref, Medline, Google Scholar
80 : Cingulate cortex of the rhesus monkey, II: cortical afferents. J Comp Neurol 1987; 262:271–289Crossref, Medline, Google Scholar
81 : Neural correlates of memories of childhood sexual abuse in women with and without posttraumatic stress disorder. Am J Psychiatry 1999; 156:1787–1795Medline, Google Scholar
82 : Smaller volume of anterior cingulate cortex in abuse-related posttraumatic stress disorder. J Affect Disord 2006; 90:171–174Crossref, Medline, Google Scholar
83 : Repeated stress induces dendritic spine loss in the rat medial prefrontal cortex. Cereb Cortex 2005; 16:313–320Crossref, Medline, Google Scholar
84 : Chronic stress induces contrasting patterns of dendritic remodeling in hippocampal and amygdaloid neurons. J Neurosci 2002; 22:6810–6818Crossref, Medline, Google Scholar
85 : The human orbitofrontal cortex: linking reward to hedonic experience. Nat Rev Neurosci 2005; 6:691–702Crossref, Medline, Google Scholar
86 : The subgenual anterior cingulate cortex in mood disorders. CNS Spectr 2008; 13:663–681Crossref, Medline, Google Scholar
87 : Forty-five years of split-brain research and still going strong. Nat Rev Neurosci 2005; 6:653–659Crossref, Medline, Google Scholar
88 : The anatomic basis of delusions after right cerebral infarction. Neurology 1984; 34:577–582Crossref, Medline, Google Scholar
89 : Possible mechanisms of anosognosia: a defect in self-awareness. Philos Trans R Soc Lond B Biol Sci 1998; 353:1903–1909Crossref, Medline, Google Scholar
90 : Neuropsychological and neuroradiologic correlates of emotional prosody comprehension. Neurology 1994; 44:515–522Crossref, Medline, Google Scholar
91 : Lack of heart rate changes during an attention-demanding task after right hemisphere lesions. Neurology 1987; 37:624–630Crossref, Medline, Google Scholar
92 : Feeling unreal: a PET study of depersonalization disorder. Am J Psychiatry 2000; 157:1782–1788Crossref, Medline, Google Scholar
93 : Brain activation during script-driven imagery induced dissociative responses in PTSD: a functional magnetic resonance imaging investigation. Biol Psychiatry 2002; 52:305–311Crossref, Medline, Google Scholar
94 : Cerebral blood flow changes in patients with conversion disorder. Psychiatry Res 1998; 83:163–168Crossref, Medline, Google Scholar
95 : Functional magnetic resonance imaging of the cerebral response to visual stimulation in medically unexplained visual loss. Psychol Med 2004; 34:583–589Crossref, Medline, Google Scholar
96 : FMRI in patients with motor conversion symptoms and controls with simulated weakness. Psychosom Med 2007; 69:961–969Crossref, Medline, Google Scholar
97 : Imaging hypnotic paralysis: implications for conversion hysteria. Lancet 2000; 355:986–987Crossref, Medline, Google Scholar
98 : Ego mechanisms of defense and personality psychopathology. J Abnorm Psychol 1994; 103:44–50Crossref, Medline, Google Scholar
99 : Alien hand syndrome. Arch Neurol 2004; 61:292–294Crossref, Medline, Google Scholar
100 : Abnormal brain activation during movement observation in patients with conversion paralysis. Neuroimage 2006; 29:1336–1343Crossref, Medline, Google Scholar

