
Disorders of the Cerebellum: Ataxia, Dysmetria of Thought, and the Cerebellar Cognitive Affective Syndrome
Abstract
Many diseases involve the cerebellum and produce ataxia, which is characterized by incoordination of balance, gait, extremity and eye movements, and dysarthria. Cerebellar lesions do not always manifest with ataxic motor syndromes, however. The cerebellar cognitive affective syndrome (CCAS) includes impairments in executive, visual-spatial, and linguistic abilities, with affective disturbance ranging from emotional blunting and depression, to disinhibition and psychotic features. The cognitive and psychiatric components of the CCAS, together with the ataxic motor disability of cerebellar disorders, are conceptualized within the dysmetria of thought hypothesis. This concept holds that a universal cerebellar transform facilitates automatic modulation of behavior around a homeostatic baseline, and the behavior being modulated is determined by the specificity of anatomic subcircuits, or loops, within the cerebrocerebellar system. Damage to the cerebellar component of the distributed neural circuit subserving sensorimotor, cognitive, and emotional processing disrupts the universal cerebellar transform, leading to the universal cerebellar impairment affecting the lesioned domain. The universal cerebellar impairment manifests as ataxia when the sensorimotor cerebellum is involved and as the CCAS when pathology is in the lateral hemisphere of the posterior cerebellum (involved in cognitive processing) or in the vermis (limbic cerebellum). Cognitive and emotional disorders may accompany cerebellar diseases or be their principal clinical presentation, and this has significance for the diagnosis and management of patients with cerebellar dysfunction.
Cerebellar disorders typically manifest with ataxia—incoordination of movement, instability of gait, impairment of articulation, and difficulty with eye movement and swallowing. It has become apparent recently that many cerebellar patients also experience changes in intellect and mood. Further, mounting evidence suggests that cerebellar pathology may be associated with alterations principally in mental function, rather than motor performance. There is a new realization that defining and understanding the role of the cerebellum in higher order behavior has the potential to benefit patients with cerebellar diseases by suggesting optimal therapeutic and rehabilitation strategies. This paper surveys the current understanding of the cerebellar role in cognitive and psychiatric disorders. To place the consideration of the cerebellum in a larger context, however, we first begin with a description of the cerebellar motor syndrome and a survey of the more common cerebellar diseases.
THE CEREBELLAR MOTOR SYNDROME
The historian Max Neuburger (1868–1955) credits the cerebellum as the structure that catapulted science into the era of the hypothesis-driven experiment.1 Thomas Willis (1621–1675) asserted that cerebellum was the seat of vegetative functions and critical for survival. This conclusion was most likely derived from the damage produced to the brainstem (medulla in particular) inflicted on awake animals by the techniques used at that time. Nicolaus Steno (Niels Stensen, 1638–1686) challenged Willis' conclusion that damage to cerebellum was incompatible with life, and performed his own experiments on cerebellum that disproved this hypothesis. In this manner, Steno's experimental disproof of Willis' hypothesis initiated the field of experimental physiology and structure—function correlation in neuroscience. Whereas Franz Josef Gall (1758–1828), arguably the founder of cognitive neuroscience, believed that cerebellum was the seat of sexual function,2 Luigi Rolando (1773–1831) concluded that it was a motor structure, and Marie-Jean-Pierre Flourens (1794–1867) determined that its role was to coordinate movements. The experimental work of Luigi Luciani,3 David Ferrier and William Aldren Turner,4 and J.S. Rissien Russell5 demonstrated motor incoordination in monkeys following cerebellar lesions. The clinical features of the now well-established cerebellar motor syndrome were defined by clinicians including Sanger Brown,6 Pierre Marie,7 Joseph François Felix Babinski,8 and Gordon Holmes.9
The cerebellar motor syndrome thus identified is characterized in contemporary terms as impairment of gait (ataxia), extremity coordination (dysmetria), disordered eye movements, poor articulation (dysarthria), impaired swallowing (dysphagia), and tremor. The basic deficit common to the motor incapacity is impairment of rate, rhythm, and force of contraction. In the early stages of cerebellar degenerative disorders, balance is poor, and there is inability to stand on one leg or perform tandem gait. As the condition progresses, walking is characterized by widened base; turning is problematic and can result in falls; and there is high stepping, staggering, and lurching from side to side. When ataxia is severe, individuals are no longer able to stand or walk without great assistance and effort. Dysmetria of the extremities is evident in dysdiadochokinesis (the impairment of alternating movements), dysrhythmic tapping of feet or hands, terminal dysmetria and swerving of the arm with finger to nose testing, side-to-side dysmetria and proximal overshoot with the heel to shin test, and decomposition of movement evident in the attempt to draw an imaginary circle in the air with the legs. The rebound phenomenon occurs (overcorrection of passive displacement of the limb), as well as overshoot of the affected extremity when following a stimulus rapidly, and sometimes tremor of extremities, head and trunk (titubation). Tone is decreased, a sign formerly thought to be the pathophysiologic basis of the motor disability. Abnormal eye movements at rest include square wave jerks, microsaccades, and chaotic movements termed opsoclonus and ocular flutter. The hallmark feature of nystagmus is present with horizontal and, less often, with vertical gaze. There are saccadic intrusions into the smooth pursuit reflex, hypometric or hypermetric saccades, slowing of saccades, impairment of the normal oculokinetic nystagmus, and loss of the ability to cancel the vestibulo-ocular reflex (as in focusing on a stationery object when the background is moving—performed with the patient rotating about an axis). Speech is characterized by “scanning” dysarthria, with alteration in rate (slower), rhythm (irregular), and force (variable volume). There is slurring of speech, tremor of the voice, and ataxic respiration. When the cerebellar motor syndrome is fully manifest it is a striking and potentially severely disabling condition. As discussed below, the cerebellar motor syndrome does not necessarily occur in all cerebellar diseases. Indeed, in monkeys, and humans with parkinsonian tremor,10 lesions of the cerebellar dentate nucleus do not produce motor disability.
DISEASES OF THE CEREBELLUM
A large number of diseases involve the cerebellum. A comprehensive consideration of these disorders is beyond the scope of this report, but many of these are listed in Table 1, and are dealt with elsewhere.11
Hereditary cerebellar degenerative diseases, in particular, have received considerable attention in recent years, as the autosomal dominant spinocerebellar ataxias (SCAs) have been identified as a distinct group of disorders that in many cases have a definable genetic basis. Margolis12 has simplified the increasingly complex set of spinocerebellar ataxias by classifying them into three discrete groups based on pathogenesis. The polyglutamine disorders, SCAs 1, 2, 3, 7, and 17, result from proteins with toxic stretches of polyglutamine. The channelopathies, SCA 6 and episodic ataxia types 1 and 2 result from disruption of potassium or calcium channel function. And the gene expression disorders, SCAs 8, 10, and 12, result from repeat expansions outside of coding regions that may quantitatively alter gene expression. The remaining SCAs 4, 5, 11, 13–16, 18–22, and 25 are presently of unknown etiology, and do not yet fit into one of these three groups.
The clinical features that potentially differentiate the genetic ataxias from each other are often unreliable, as they may be inconsistent in a given SCA, or are shared by more than one of the SCAs. This is further compounded by the recognition that Friedreich's ataxia, the autosomal recessive GAA triplet repeat disorder on chromosome 9, was formerly considered to be a disease of childhood with an unmistakable phenotype of ataxia, areflexia, extensor plantar responses, neuropathy, scoliosis and cardiomyopathy. Friedreich's ataxia is now know to present well into adulthood (late-onset Friedreich's ataxia [LOFAR]) or have a phenotype marked only by ataxia and preserved or brisk deep tendon reflexes (Friedreich's ataxia with retained reflexes [FARR]). Moreover, the diagnostic challenge to the clinician is complicated by the knowledge that cerebellar features may be principal manifestations of a number of other system diseases, including mitochondrial disorders.
NONMOTOR ASPECTS OF CEREBELLAR FUNCTION
Based on anatomic, physiologic, and clinical information, investigators such as Ray Snider,13 Robert Dow,14 and Robert Heath15 suggested that the cerebellum is a great modulator of neurologic function, and that whatever it does to motor control, it also does the same thing to other kinds of behaviors. Watson16 discussed the relevance of the cerebellum to higher order function, Frick17 explored how the vestibulocerebellum was related to the ego, and S.R. Snyder18 addressed its role in schizophrenia. The incorporation of the cerebellum into the distributed cortico-subcortical neural system subserving cognition was more formally developed as a concept by the Leiners with Dow19 based on the expansion of the neocerebellum and dentate nucleus along with the cerebral association areas; by Schmahmann and Pandya20 and Schmahmann21 based on earlier physiology and clinical observations, and the associative corticopontine projections to the basilar pons in the monkey; by Strick et al.22 based on the feedback from cerebellar neodentate nucleus to the prefrontal cortex; and by a number of subsequent investigators.
Since the earliest days of investigation into cerebellar function, the possibility that the cerebellum is involved in areas of neurological processing beyond motor control has been raised explicitly. Around the time that Flourens concluded that cerebellum is a motor control device, and long before the work of the behaviorists and clinicians that firmly entrenched the notion of cerebellum as a motor apparatus, investigators (perhaps starting with Combettes23) began to report that cerebellar pathology was associated with clinical manifestations outside of the motor domain. Reports throughout the 1800s described individuals with different forms of cerebellar damage, including failure of development (agenesis) and cerebellar atrophy, who demonstrated impairments of intellectual function and emotional or psychiatric disturbances.21 Later clinical studies identified a relationship between the cerebellum and personality, aggression and emotion,24 and they linked psychosis—schizophrenia in particular—with enlargement of the fourth ventricle, smaller cerebellar vermis, and cerebellar atrophy.25,26 In the middle and later decades of the 20th century, before the genetic basis of the ataxias was identified and multiple systems atrophy was classified with the synucleinopathies,27 patients with cerebellar cortical atrophy and what was known as olivopontocerebellar atrophy were found to have cognitive problems. These included impairments in verbal intelligence, visual-spatial abilities, learning and memory, and frontal system functions.28–31 Tests showed deficits in strategy formation32 and procedural learning.33
Now that genetic diagnosis of many of the spinocerebellar ataxias is available, new information suggests that there is some degree of cognitive change at different stages in most of the SCAs,34 although the pattern of neuropsychological deficits has not yet been shown to distinguish between these disorders. Impaired executive functions, deficits in verbal short-term memory, and mild, generalized cognitive impairment35 have been documented in SCA 1 (CAG repeat on chromosome 6). Patients with SCA 2 (triplet repeat on chromosome 12) may develop poor memory, concentration problems, impairments of conceptual reasoning, and frontal-executive dysfunction on tests of verbal fluency and Luria's test of motor set switching, as well as emotional instability and impulsivity.36,37 Individuals with Machado-Joseph disease or SCA 3 (CAG expansion on chromosome 14) demonstrate deficits in visual and verbal attention, verbal fluency, and planning and strategy tests such as the Wisconsin Card Sort Test.38
In SCA 13 (chromosome 19), moderate mental retardation (IQ 62–76) occurs in association with a progressive childhood-onset cerebellar “motor” syndrome and developmental delay.39 Dementia is part of the diagnosis in SCA 17 (dementia, psychosis, extrapyramidal feature and seizures, with a CAG repeat on chromosome 6,40,41) and in SCA 21 (ataxia with hyporeflexia, akinesia, rigidity, and cognitive impairment in a French kindred, with linkage to chromosome 7).42 In Friedreich's ataxia the major pathology is located outside cerebellum and the extent of neuropsychological impairment has been studied with varying results. Some have noted impairments on tests of visual-perceptual and visual-constructive abilities, slowed information processing speed, decreased sustained attention, reduced verbal span, deficits in letter fluency, and impaired acquisition and consolidation of verbal information.43 In contrast, other studies have shown patients with Friedreich's ataxia to be relatively cognitively intact.44
Pathological features are not confined to the cerebellum in most of these hereditary ataxias, and so the cognitive and emotional impairments are unlikely to be related exclusively to cerebellum.
In patients with focal cerebellar lesions, language problems include agrammatism,45 decreased verbal fluency,46 and inability to detect one's own errors in tasks such as the verb-for-noun generation paradigm.47 Patients with ataxia have difficulties with attentional modulation,48 motor skill learning,49–51 and the ability to acquire conditional associative reflexes.52 Experimental studies in animals reveal that electrical potentials are activated in cerebellum by stimulation of somatosensory, visual, and auditory cortices;53 stimulation of deep cerebellar nuclei evokes autonomic responses,54,55 grooming, predatory attack, and sham rage;56,57 and lesions of the cerebellar anterior interpositus nucleus prevent or abolish conditional associative learning tasks.58
This clinical and experimental background notwithstanding, a persistent set of clinical questions has limited the consideration of the cerebellum as an integral component of the circuits that subserve cognition and emotion. That is, are the reported cognitive impairments in cerebellar patients observed only with subtle neuropsychological tests, or are they indeed relevant for patients' lives? Do the deficits result from cerebellar damage itself, or from lesions in other brain regions affected by the neurodegenerative disorders?
THE CEREBELLAR COGNITIVE AFFECTIVE SYNDROME
Adults
In order to address the clinical uncertainties regarding the effect of focal cerebellar lesions on higher order behaviors, we performed neurological examinations, bedside mental state tests and neuropsychological testing in 20 patients with lesions confined to the cerebellum.59 Thirteen patients had stroke, three postinfectious cerebellitis, three cerebellar cortical atrophy, and one excision of a midline tumor. We determined that there was a pattern of clinically relevant behavioral changes that could be diagnosed at the bedside and further quantified by neuropsychological tests. We termed this the “cerebellar cognitive affective syndrome” (CCAS). It is characterized by 1) disturbances of executive function, which includes deficient planning, set-shifting, abstract reasoning, working memory, and decreased verbal fluency; 2) impaired spatial cognition, including visual-spatial disorganization and impaired visual-spatial memory; 3) personality change characterized by flattening or blunting of affect and disinhibited or inappropriate behavior; and 4) linguistic difficulties, including dysprosodia, agrammatism and mild anomia. The net effect of these disturbances in cognitive functioning was a general lowering of overall intellectual function (Table 2).
These impairments were present on routine bedside mental state tests, and confirmed using standardized neuropsychological tests. They were clinically relevant, were noted by family members and the medical staff, and could not be explained by difficulties with motor control because in many cases the motor incoordination was mild. The neurobehavioral presentation was more pronounced and generalized in those with bilateral or large unilateral infarctions in the posterior lobes in the territory of the posterior inferior cerebellar arteries, and in those with subacute onset of pancerebellar disorders such as occurs with postinfectious cerebellitis. It was less evident in patients with more slowly progressive cerebellar degenerations, in the recovery phase (3–4 months) after acute stroke and in those with restricted cerebellar pathology (smaller strokes in the territory supplied by the superior cerebellar artery, that is, in the anterior lobe of the cerebellum or the rostral part of the posterior lobe). The vermis was consistently involved in patients with pronounced affective presentations.
The clinical relevance of the CCAS has been replicated in subsequent reports of adults with cerebellar strokes. Malm et al.60 demonstrated deficits in attention, working memory, visuospatial skills, and cognitive flexibility Stroke patients studied by Neau and others61 had deficits in executive function, spatial cognition, attention, and some language tasks. Single case reports have shown that patients with right cerebellar infarction develop impairments of linguistic processing including agrammatism and decreased verbal fluency.46
Children
An underrecognized but important aspect of cerebellar function is its impact on cognition and emotion in children with cerebellar disorders. Levisohn et al.62 documented that the CCAS occurs in children as well. We found that there were characteristic behavioral deficits in 19 children ages 3 to 14 who had undergone resection of cerebellar tumors. These included difficulties with initiation of language, impaired verbal fluency and word finding difficulties, decreased story retrieval, deficits in sequencing, planning and maintaining set, and visual-spatial deficits. Impaired regulation of affect was seen in children with damage to the vermis, including irritability, impulsivity, disinhibition, and lability of affect with poor attentional and behavioral modulation. Riva and Giorgi63 observed similar phenomena in their series of children following resection of cerebellar tumors, namely, impairment in verbal intelligence and complex language tasks following right cerebellar hemisphere lesions, and deficient nonverbal tasks and prosody after left cerebellar hemisphere lesions. Children with vermal involvement developed irritability and autistic-like features.
Five children in our study exhibited the posterior fossa syndrome. This phenomenon is observed in approximately 15% of children who undergo resection of midline cerebellar tumors,64,65 although there was a higher incidence in our study. It is characterized by the development of mutism 1 to 4 days postoperatively, and in the recovery phase over a period of months it is accompanied by dysarthria, buccal and lingual apraxia, and a behavioral syndrome that includes regressive personality changes, apathy, withdrawal and poverty of spontaneous movement. Emotional lability is marked, and there is rapid fluctuation of expression of emotion that gravitates between irritability with inconsolable crying and agitation, to giggling and easy distractibility.66
Developmental Anomalies
Cognitive and behavioral problems in children with cerebellar lesions have been observed in the setting of absence of the cerebellum (either complete or partial agenesis), as well as nonprogressive cerebellar ataxia—what used to be called ataxic cerebral palsy. It used to be taught that cerebellar agenesis is asymptomatic, but this appears to be incorrect, both from the perspective of motor symptoms67 as well as higher order behavior. Gardner et al.68 reported delayed milestones, mild motor impairments, and intellectual handicap in three patients with near-near total absence of the cerebellum. We found69 that near-complete or partial cerebellar agenesis in six children was accompanied by behavioral and motor deficits. The severity and range of the motor, cognitive, and psychiatric impairments were greater in those with more pronounced agenesis. The children presented with gross and fine motor delay, oral motor apraxia, impaired saccades and vestibulo-ocular reflex cancellation, clumsiness, and mild ataxia. Behavioral features included autistic-like stereotypical performance, obsessive rituals, and difficulty understanding social cues. Tactile defensiveness (avoidance of and adverse reaction to touch), was a prominent feature in four children. Executive impairments included perseveration, disinhibition, and poor abstract reasoning, working memory and verbal fluency. Spatial cognition was impaired for perceptual organization, visual spatial copying and recall. Some children presented with expressive language delay as the principal manifestation, in two instances so severe as to require instruction in sign language. Impaired prosody was evident in all cases, and overregularization of past tense verbs was noted. In longitudinal follow up, extensive rehabilitation enhanced motor, linguistic, and cognitive performance.
Several observations suggest a relationship between developmental anomalies of cerebellum and neurobehavioral syndromes. Quantitative morphometry of the cerebellum in attention deficit hyperactivity disorder reveals smaller posterior lobes of the vermis in both males70,71 and females,72 and the size of the vermis seems to be related to the severity of this disorder. Allin et al.73 found that the cerebellum is significantly smaller in children who were born very preterm (before 33 weeks of gestation), and this is correlated with impaired executive and visual-spatial functions, as well as impaired language skills—the principal features of the CCAS. One of the theories of the pathophysiology of dyslexia is that cerebellar abnormalities prevent normal eye movements and interfere with the acquisition of lexical information.74 Nicolson et al.75 found support for this hypothesis by showing that individuals with dyslexia have lower cerebellar activation on positron emission tomography scans compared to control subjects when learning novel sequences and when executing prelearned sequences of finger movements.
Psychiatric Disorders
Adults and children with the CCAS may experience altered regulation of mood and personality, display obsessive-compulsive tendencies, and demonstrate psychotic thinking. As discussed above, early accounts of the behavioral consequences of cerebellar lesions focused on psychiatric manifestations and the morphologic changes in cerebellum in schizophrenia and other psychoses. The notion that there is a cerebellar component to the pathophysiology of schizophrenia is gaining momentum from current morphometric, metabolic, and functional imaging studies. Nopoulos et al.76 performed automated volumetric measures of subregions of the vermis in patients with schizophrenia and found that the anterior lobe vermis was smaller than in control subjects, and its size was positively correlated with total cerebellar volume, temporal lobe volume, and full-scale IQ. Further, lower cerebellar volume in patients with schizophrenia was associated with the duration of negative and psychotic symptoms and with psychosocial impairment.77 The vermis was decreased in size despite overall increased cerebellar blood volume in the schizophrenic patients studied by Loeber et al.78 These investigators79 also found that compared with control subjects, patients with schizophrenia have a significantly smaller inferior vermis and less cerebellar hemispheric asymmetry. Similarly, Ichimiya et al.80 found significantly reduced vermis volume in neuroleptic-naive schizophrenic patients that correlated with the Depression and Paranoia subscores of the Brief Psychiatric Rating Scale and Volz et al.81 found reduced volumes in the left cerebellar hemisphere and right cerebellar vermis, along with smaller volumes in frontal lobe, temporal lobe, and thalamus. Not all studies demonstrate smaller cerebellar volumes.82 Levitt et al.83 observed that vermis volume was greater in their schizophrenia population, vermis white matter volume correlated with severity of positive symptoms, thought disorder, and impairment in verbal logical memory, and patients showed a trend for more cerebellar hemispheric volume asymmetry (left greater than right).
Metabolic abnormalities on magnetic resonance spectroscopy of chronically medicated male patients with schizophrenia include decreased N-acetylaspartate, a putative neuronal/axonal marker, and creatine in the anterior cerebellar vermis, independent of duration of illness or neuroleptic dose.84 Positron emission tomography (PET) in neuroleptic-naive patients with schizophrenia85 has revealed increased metabolic activity in cerebellum, thalamus and retrosplenial cortex, along with decreased metabolic activity in prefrontal, inferotemporal and parietal cortices. Schizophrenic patients perform tests of word list recall at similar levels to control subjects but have decreased PET activation in the cerebellum, as well as in frontal and temporal lobe areas and thalamus.86
Individuals with early infantile autism, once classified as juvenile schizophrenia, have enlarged IV ventricles, loss of Purkinje cells in the lateral and inferior cerebellar cortex, and abnormal or reduced numbers of neurons in the deep cerebellar nuclei.87–90 Vermal91 and hemispheric92 atrophy are evident on some magnetic resonance imaging (MRI) studies of autistic subjects, although these findings remain controversial.93 Pathological findings are present in other brain regions in autism, particularly in the limbic system, and thus it remains to be established which anatomic-pathologic features are most closely associated with the autistic features, and whether these pathologic findings cause the behavioral aberrations.
Depression has been shown to be a recurring problem in patients with spinocerebellar ataxia. Leroi et al.94 found that more than half their patients with cerebellar degeneration had psychopathology including depression, personality change, psychotic disorders, and impaired cognition. Affected individuals endorsed symptoms of depression that reach moderate-to-severe levels, as well as apathy. In another study,95 cerebellar vermal lobules VIII through X were smaller in patients with bipolar disorder who experienced multiple episodes of depression. This suggested that cerebellar vermal atrophy may be a late neurodegenerative event in those who have multiple affective episodes. It is important that depression be recognized in cerebellar patients, because it can be treated.
The mechanisms of the cerebellar modulation of complex behavior remain to be established. Case studies in patients with emotional dysregulation have begun to address this. Parvizi et al.96 explored the dysequilibrium inherent in the emotional display of some patients with cerebellar lesions by considering the phenomenon of pathologic laughter and crying in a patient in whom the cerebellum was partially deafferented by multiple infarcts. The authors concluded that pathologic laughter and crying arose from disruption of the cortico-ponto-cerebellar pathways, preventing the cerebellum from automatically adjusting the execution of emotional display to cognitive and situational context, and resulting in inadequate or chaotic behavior. Annoni et al.97 were impressed by emotional blunting in a patient who underwent surgical excision of an infarcted left cerebellum (posterior and anterior inferior cerebellar artery territories). In addition to the loss of emotional display and concern, testing revealed other aspects of the CCAS including impaired cognitive flexibility and decision making ability, as well as increased risk taking on the gambling task of Bechara et al.98 Autonomic reactions assessed by skin conductance to positively and negatively charged stimuli were undifferentiated, suggesting that the healthy cerebellum is involved in the generation of emotionally congruent autonomic reactions. Further analysis of the mechanisms subserving the role of the cerebellum in the modulation of autonomic responses and complex behavior will be of great interest.
FUNCTIONAL IMAGING
Functional brain imaging has been very informative, and has revealed that cerebellum is activated by a large number of cognitive tasks that are devoid of movement. This large field cannot be summarized here, but it is worth noting that nonmotor domains that invoke a cerebellar contribution include sensory processing and discrimination, mental imagery, motor learning, classical conditioning, nonmotor learning and memory, linguistic processing, attentional modulation, timing estimation, emotion perception and experience, visual spatial memory, executive function (including verbal working memory, strategy, reasoning, and verbal fluency), and autonomic functions including the experience and anticipation of pain, thirst, hunger, and smell. There is evidence to suggest that these functions are under the control of different areas within the cerebellum.99–101
DYSMETRIA OF THOUGHT AND THE UNIVERSAL CEREBELLAR TRANSFORM
The attempt to understand how cerebellum may be involved in all these higher order brain functions is facilitated by a consideration of the anatomy of the cerebellum and its connections with the cerebral hemispheres and the brainstem. The microscopic anatomy of the cerebellar cortex is quite uniform,102 and it may therefore seem puzzling that the cerebellum could be involved in all these different aspects of neurologic and psychological function. Anatomic tract tracing studies indicate that there are pathways linking the cerebellum with autonomic,103,104 limbic,105 and associative regions of the cerebral cortex106,107 as well as with sensorimotor cortices. This allows the cerebellum to communicate with brain areas concerned with instinctive behaviors, mood, and the highest levels of cognition and reasoning. Together with Deepak Pandya, we extended the work of earlier anatomists by demonstrating that the association areas in the prefrontal cortices, posterior parietal lobes, superior temporal regions, and parahippocampal areas, send information in a precisely organized manner to the nuclei of the basilar pons,20,106–110 from where the information is conveyed to the cerebellum. Middleton and Strick22 demonstrated that the deep cerebellar nuclei, notably the dentate, send information back to those areas of the prefrontal cortex that send information into the cerebellum. Further, just as there is a precise ordering in the way that information is sent from the cerebral cortex into the pons, and from there to the cerebellum, the feedback from the dentate nucleus of the cerebellum to the cerebral cortex is also precisely arranged. Thus, there are circuits, or anatomic loops that link higher order areas of the brain with the cerebellum, in a bidirectional manner. (This anatomical work is summarized in the 1997 monograph, The Cerebellum and Cognition.)100
It has become necessary to develop a new way of thinking about the cerebellum, one that takes all the various cerebellar roles into consideration. If cerebellum is not only a motor control device, then what does it do, and how does it do it? The early notion that the role of the cerebellum is to modulate neurologic function is compelling, and we have adopted and amended this as part of a conceptual approach to cerebellar function that I have referred to as the “dysmetria of thought hypothesis.”21,111,112 In this view, because cerebellar anatomy is essentially uniform throughout the structure, the basic work that cerebellum does in the nervous system should be constant as well. This we have referred to as the universal cerebellar transform, characterized as the cerebellar modulation of behavior, serving as an oscillation dampener maintaining function automatically around a homeostatic baseline and smoothing out performance in all domains (Table 3).
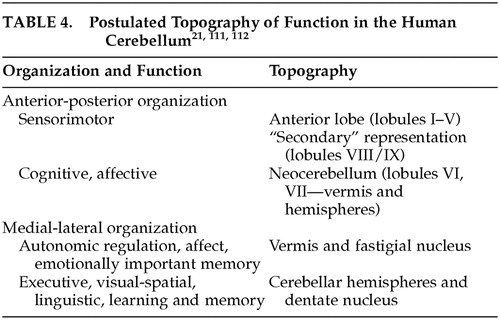
The specificity of the anatomic subcircuits in the cerebrocerebellar system indicates that different areas in the cerebellum interact with precise and different areas of the cerebral cortex. These anatomic subcircuits are the structural basis for putative functional subunits, and facilitate what appears to be topographic organization of motor and cognitive function in the cerebellum. In this proposed schema, the anterior lobe is mainly involved with motor control, whereas the posterior lobe is more concerned with higher order behaviors. Further, whereas the lateral parts of the posterior lobe are thought to be involved in cognitive operations, the vermis is considered to be the equivalent of the limbic cerebellum (Table 4).
According to this hypothesis, the universal cerebellar transform is the essential functional contribution that the cerebellum makes to the distributed neural system. By corollary, therefore, there should be a universal cerebellar impairment. This universal cerebellar impairment, the hypothesis holds, is dysmetria. When the dysmetria involves the motor domain the various manifestations of ataxia are evident in extremity movements, eye movements, speech and equilibrium. However, when the dysmetria involves nonmotor functions subserved by cerebellum, this results in dysmetria of thought, or cognitive dysmetria, and manifests as the various components of the cerebellar cognitive affective syndrome.
CONCLUSIONS
Patients with cerebellar dysfunction may struggle with depression and other forms of psychological distress, limitations in cognitive ability and flexibility, slowed reaction times and impaired attentional modulation, as well as less ability to do “multitasking” automatically. These important aspects of higher order behavior have an impact on quality of life, employment, and personal relationships and need to be recognized by the medical profession as well as by patients and their families. By working with available treatments and novel cognitive rehabilitation strategies, adults and children with inherited or acquired cerebellar disorders could benefit from the new recognition that the cerebellum is not only a motor control device, but it is also an essential component of the brain mechanisms for personality, mood, and intellect.
ACKNOWLEDGMENTS
This study was supported in part by NIMH grant 1R01 MH-067980-1 and the Birmingham Foundation. The assistance of Jason MacMore is gratefully acknowledged.
 |
 |

1 Neuburger M, Clarke E: The Historical Development of Experimental Brain and Spinal Cord Physiology Before Flourens (1868). Translated and edited by Clarke E. Baltimore, Johns Hopkins University Press, 1981Google Scholar
2 Clarke E, O'Malley CD: The Human Brain and Spinal Cord: A Historical Study Illustrated by Writings From Antiquity to the Twentieth Century, 2nd ed. San Francisco, Norman Publishing, 1996Google Scholar
3 Luciani L: Il cervelletto: Nuovi studi di fisiologia normale e patologica. Firenze, Italy, Le Monnier, 1891Google Scholar
4 Ferrier D, Turner WA: A record of experiments illustrative of the symptomatology and degenerations following lesions of the cerebellum and its peduncles and related structures in monkeys. Phil Trans R Soc Lond B 1893; 185:719–778Crossref, Google Scholar
5 Russell JSR: Experimental researches into the functions of the cerebellum. Phil Trans R Soc Lond B 1894; 185:819–861Crossref, Google Scholar
6 Brown S: On hereditary ataxia, with a series of twenty-one cases. Brain 1892; 15:250–282Crossref, Google Scholar
7 Marie P: Sur l'hérédo-ataxie cérébelleuse. Semaine méd 1893; 13:444–447Google Scholar
8 Babinski JFF: De l'asynergie cerebelleuse. Rev Neurol 1899; 7:806–816Google Scholar
9 Holmes G: The symptoms of acute cerebellar injuries due to gunshot wounds. Brain 1917; 40:461–535Crossref, Google Scholar
10 Zervas NT, Horner FG, Gordy PD: Cerebellar dentatectomy in primates and humans. Trans Am Neurol Assoc 1967; 92:27–30Medline, Google Scholar
11 Manto M, Pandolfo M: The Cerebellum and Its Disorders. New York, Cambridge University Press, 2001Google Scholar
12 Margolis RL: The spinocerebellar ataxias: order emerges from chaos. Curr Neurol Neurosci Rep 2002; 2:447–456Crossref, Medline, Google Scholar
13 Snider RS, Maiti A: Cerebellar contributions to the Papez circuit. J Neurosci Res 1976;2:133–146Google Scholar
14 Dow RS: Some novel concepts of cerebellar physiology. Mt Sinai J Med 1974; 41:103–119Medline, Google Scholar
15 Heath RG: Modulation of emotion with a brain pacemaker: treatment for intractable psychiatric illness. J Nerv Ment Dis 1977; 165:300–317Crossref, Medline, Google Scholar
16 Watson PJ: Nonmotor functions of the cerebellum. Psychol Bull 1978; 85:944–967Crossref, Medline, Google Scholar
17 Frick RB: The ego and the vestibulocerebellar system. Psychoanal Q 1982; 51:93–122Crossref, Medline, Google Scholar
18 Snider SR: Cerebellar pathology in schizophrenia—cause or consequence? Neurosci Behav Rev 1982;6:47–53Google Scholar
19 Leiner HC, Leiner AL, Dow RS: Does the cerebellum contribute to mental skills? Behav Neurosci 1986; 100:443–454Crossref, Medline, Google Scholar
20 Schmahmann JD, Pandya DN: Anatomical investigation of projections to the basis pontis from posterior parietal association cortices in rhesus monkey. J Comp Neurol 1989; 289:53–73Crossref, Medline, Google Scholar
21 Schmahmann JD: An emerging concept: the cerebellar contribution to higher function. Archiv Neurol 1991; 48:1178–1187Crossref, Medline, Google Scholar
22 Middleton FA, Strick PL: Anatomical evidence for cerebellar and basal ganglia involvement in higher cognitive function. Science 1994; 266:458–461Crossref, Medline, Google Scholar
23 Combettes: Absence complete du cervelet, des pedoncles posterieurs et la protuberance cerebrale chez une jeune fille morte dans sa onzieme année. Bull Soc Anat de Paris 1831; 5:148–157Google Scholar
24 Heath RG, Franklin DE, Shraberg D: Gross pathology of the cerebellum in patients diagnosed and treated as functional psychiatric disorders. J Nerv Ment Dis 1979; 167:585–592Crossref, Medline, Google Scholar
25 Moriguchi I: A study of schizophrenic brains by computerized tomography scans. Folia Psychiatr Neurol Jpn 1981; 35:55–72Medline, Google Scholar
26 Lippmann S, Manshadi M, Baldwin H, et al: Cerebellar vermis dimensions on computerized tomographic scans of schizophrenic and bipolar patients. Am J Psychiatry 1982; 139:667–668Crossref, Medline, Google Scholar
27 Dickson DW, Lin W, Liu WK, et al: Multiple system atrophy: a sporadic synucleinopathy. Brain Pathol 1999; 9:721–732Crossref, Medline, Google Scholar
28 Landis DMD, Rosenberg RN, Landis SC, et al: Olivopontocerebellar degeneration. Arch Neurol 1974; 31:295–307Crossref, Medline, Google Scholar
29 Kish SJ, El-Awar M, Schut L, et al: Cognitive deficits in olivopontocerebellar atrophy: implications for the cholinergic hypothesis of Alzheimer's dementia. Ann Neurol 1988; 24:200–206Crossref, Medline, Google Scholar
30 Bracke-Tolkmitt R, Linden A, Canavan AGM, et al: The cerebellum contributes to mental skills. Behav Neurosci 1989; 103:442–446Crossref, Google Scholar
31 Botez-Marquard T, Botez MI: Cognitive behavior in heredodegenerative ataxias. Eur Neurol 1993; 33:351–357Crossref, Medline, Google Scholar
32 Grafman J, Litvan I, Massaquoi S, et al: Cognitive planning deficit in patients with cerebellar atrophy. Neurology 1992; 42:1493–1496Crossref, Medline, Google Scholar
33 Pascual-Leone A, Grafman J, Clark K, et al: Procedural learning in Parkinson's disease and cerebellar degeneration. Ann Neurol 1993; 34:594–602Crossref, Medline, Google Scholar
34 Geschwind DH: Focusing attention on cognitive impairment in spinocerebellar ataxia. Arch Neurol 1999; 56:20–22Crossref, Medline, Google Scholar
35 Burk K, Bosch S, Globas C, et al: Executive dysfunction in spinocerebellar ataxia type 1. Eur Neurol 2001; 46:43–48Crossref, Medline, Google Scholar
36 Gambardella A, Annesi G, Bono F, et al: CAG repeat length and clinical features in three Italian families with spinocerebellar ataxia type 2 (SCA2): early impairment of Wisconsin Card Sorting Test and saccade velocity. J Neurol 1998; 245:647–652Crossref, Medline, Google Scholar
37 Storey E, Forrest SM, Shaw JH, et al: Spinocerebellar ataxia type 2: clinical features of a pedigree displaying prominent frontal-executive dysfunction. Arch Neurol 1999; 56:43–50Crossref, Medline, Google Scholar
38 Zawacki TM, Grace J, Friedman JH, et al: Executive and emotional dysfunction in Machado-Joseph disease. Mov Disord 2002; 17:1004–1010Crossref, Medline, Google Scholar
39 Herman-Bert A, Stevanin G, Netter JC, et al: Mapping of spinocerebellar ataxia 13 to chromosome 19q13.3-q13.4 in a family with autosomal dominant cerebellar ataxia and mental retardation. Am J Hum Genet 2000; 67:229–235Crossref, Medline, Google Scholar
40 Koide R, Kobayashi S, Shimohata T, et al: A neurological disease caused by an expanded CAG trinucleotide repeat in the TATA-binding protein gene: a new polyglutamine disease? Hum Molec Genet 1999; 8:2047–2053Crossref, Medline, Google Scholar
41 Nakamura K, Jeong S-Y, Uchihara T, et al: SCA17, a novel autosomal dominant cerebellar ataxia caused by an expanded polyglutamine in TATA-binding protein. Hum Molec Genet 2001; 10:1441–1448Crossref, Medline, Google Scholar
42 Devos D, Schraen-Maschke S, Vuillaume I, et al: Clinical features and genetic analysis of a new form of spinocerebellar ataxia. Neurology 2001; 56:234–238Crossref, Medline, Google Scholar
43 Wollmann T, Barroso J, Monton F, et al: Neuropsychological test performance of patients with Friedreich's ataxia. J Clin Exp Neuropsychol 2002; 24:677–686Crossref, Medline, Google Scholar
44 White M, Lalonde R, Botez-Marquard T: Neuropsychologic and neuropsychiatric characteristics of patients with Friedreich's ataxia. Acta Neurol Scand 2000; 102:222–226Crossref, Medline, Google Scholar
45 Silveri MC, Leggio MG, Molinari M: The cerebellum contributes to linguistic production: a case of agrammatic speech following a right cerebellar lesion. Neurology 1994; 44:2047–2050Crossref, Medline, Google Scholar
46 Molinari M, Leggio MG, Silveri MC: Verbal fluency and agrammatism. Int Rev Neurobiol 1997; 41:325–339Crossref, Medline, Google Scholar
47 Fiez JA, Petersen SE, Cheney MK, et al: Impaired non-motor learning and error detection associated with cerebellar damage. Brain 1992; 115:155–178Crossref, Medline, Google Scholar
48 Courchesne E, Townsend J, Akshoomoff NA, et al: Impairment in shifting attention in autistic and cerebellar patients. Behav Neurosci 1994; 108:848–865Crossref, Medline, Google Scholar
49 Sanes JN, Dimitrov B, Hallett M: Motor learning in patients with cerebellar dysfunction. Brain 1990; 113:103–120Crossref, Medline, Google Scholar
50 Thach WT, Goodkin HP, Keating JG: The cerebellum and the adaptive coordination of movement. Annu Rev Neurosci 1992; 15:403–442Crossref, Medline, Google Scholar
51 Doyon J, Song AW, Karni A, et al: Experience-dependent changes in cerebellar contributions to motor sequence learning. Proc Natl Acad Sci USA 2002; 99:1017–1022Crossref, Medline, Google Scholar
52 Solomon PR, Stowe GT, Pendlebury WW: Disrupted eyelid conditioning in a patient with damage to cerebellar afferents. Behav Neurosci 1989; 103:898–902Crossref, Medline, Google Scholar
53 Snider RS: Recent contributions to the anatomy and physiology of the cerebellum. AMA Arch Neurol Psychiatry 1950; 64:196–219Crossref, Medline, Google Scholar
54 Zanchetti A, Zoccolini A: Autonomic hypothalamic outbursts elicited by cerebellar stimulation: J Neurophysiol 1954; 17:473–483Google Scholar
55 Martner J: Cerebellar influences on autonomic mechanisms. Acta Physiol Scand Suppl 1975; 425:1–42Medline, Google Scholar
56 Moruzzi G: Sham rage and localized autonomic responses elicited by cerebellar stimulation in the acute thalamic cat. Proceedings of the XVII International Congress on Physiology, Oxford, 1947, pp 114–115Google Scholar
57 Reis DJ, Doba N, Nathan MA: Predatory attack, grooming and consummatory behaviors evoked by electrical stimulation of cat cerebellar nuclei. Science 1973; 182:845–847Crossref, Medline, Google Scholar
58 Thompson RF, Bao S, Chen L, et al: Associative learning. Int Rev Neurobiol 1997; 41:151–189Crossref, Medline, Google Scholar
59 Schmahmann JD, Sherman JC: The cerebellar cognitive affective syndrome. Brain 1998; 121:561–579Crossref, Medline, Google Scholar
60 Malm J, Kristensen B, Karlsson T, et al: Cognitive impairment in young adults with infratentorial infarcts. Neurology 1998; 51: 433–440Google Scholar
61 Neau JP, Arroyo-Anllo E, Bonnaud V, et al: Neuropsychological disturbances in cerebellar infarcts. Acta Neurol Scand 2000; 102:363–370Crossref, Medline, Google Scholar
62 Levisohn L, Cronin-Golomb A, Schmahmann JD: Neuropsychological consequences of cerebellar tumor resection in children: cerebellar cognitive affective syndrome in a pediatric population. Brain 2000; 123:1041–1050Crossref, Medline, Google Scholar
63 Riva D, Giorgi C: The cerebellum contributes to higher function during development: evidence from a series of children surgically treated for posterior fossa tumors. Brain 2000; 123:1051–1061Crossref, Medline, Google Scholar
64 Wisoff JH, Epstein FJ: Pseudobulbar palsy after posterior fossa operation in children. Neurosurgery 1984; 15:707–709Crossref, Medline, Google Scholar
65 Kingma A, Mooij JJA, Metemaekers JDM, et al: Transient mutism and speech disorders after posterior fossa surgery in children with brain tumors. Acta Neurochir 1994; 131:74–79Crossref, Medline, Google Scholar
66 Pollack IF, Polinko P, Albright AL, et al: Mutism and pseudobulbar symptoms after resection of posterior fossa tumors in children: Incidence and pathophysiology. Neurosurgery 1995; 37:885–893Crossref, Medline, Google Scholar
67 Glickstein M: Cerebellar agenesis. Brain 1994; 117:1209–1212Crossref, Medline, Google Scholar
68 Gardner RJ, Coleman LT, Mitchell LA, et al: Near-total absence of the cerebellum. Neuropediatrics 2001; 32:62–68Crossref, Medline, Google Scholar
69 Chheda M, Sherman J, Schmahmann JD: Neurologic, psychiatric and cognitive manifestations in cerebellar agenesis. Neurology 2002; 58(suppl 3):356Google Scholar
70 Berquin PC, Giedd JN, Jacobsen LK, et al: Cerebellum in attention-deficit hyperactivity disorder: a morphometric MRI study. Neurology 1998; 50:1087–1093Crossref, Medline, Google Scholar
71 Mostofsky SH, Reiss AL, Lockhart P, et al: Decreased cerebellar posterior vermis size in fragile X syndrome: correlation with neurocognitive performance. Neurology 1998; 50:121–130Crossref, Medline, Google Scholar
72 Castellanos FX, Giedd JN, Berquin PC, et al: Quantitative brain magnetic resonance imaging in girls with attention-deficit/hyperactivity disorder. Arch Gen Psychiatry 2001; 58:289–295Crossref, Medline, Google Scholar
73 Allin M, Matsumoto H, Santhouse AM, et al: Cognitive and motor function and the size of the cerebellum in adolescents born very pre-term. Brain 2001; 124:60–66Crossref, Medline, Google Scholar
74 Nicolson RI, Fawcett AJ, Dean P: Developmental dyslexia: the cerebellar deficit hypothesis. Trends Neurosci 201; 24:508–511Google Scholar
75 Nicolson RI, Fawcett AJ, Berry EL, et al: Association of abnormal cerebellar activation with motor learning difficulties in dyslexic adults. Lancet 1999; 353:1662–1667Crossref, Medline, Google Scholar
76 Nopoulos PC, Ceilley JW, Gailis EA, et al: An MRI study of cerebellar vermis morphology in patients with schizophrenia: evidence in support of the cognitive dysmetria concept. Biol Psychiatry 1999; 46:703–711Crossref, Medline, Google Scholar
77 Wassink TH, Andreasen NC, Nopoulos P, et al: Cerebellar morphology as a predictor of symptom and psychosocial outcome in schizophrenia. Biol Psychiatry 1999; 45:41–48Crossref, Medline, Google Scholar
78 Loeber RT, Sherwood AR, Renshaw PF, et al: Differences in cerebellar blood volume in schizophrenia and bipolar disorder. Schizophr Res 1999; 37:81–89Crossref, Medline, Google Scholar
79 Loeber RT, Cintron CMB, Yurgelun-Todd DA: Morphometry of individual cerebellar lobules in schizophrenia. Am J Psychiatry 2001; 158:952–954Crossref, Medline, Google Scholar
80 Ichimiya T, Okubo Y, Suhara T, et al: Reduced volume of the cerebellar vermis in neuroleptic-naive schizophrenia. Biol Psychiatry 2001; 49:20–27Crossref, Medline, Google Scholar
81 Volz H, Gaser C, Sauer H: Supporting evidence for the model of cognitive dysmetria in schizophrenia—a structural magnetic resonance imaging study using deformation-based morphometry. Schizophr Res 2000; 46:45–56Crossref, Medline, Google Scholar
82 Staal WG, Hulshoff Pol HE, Schnack HG, et al: Structural brain abnormalities in chronic schizophrenia at the extremes of the outcome spectrum. Am J Psychiatry 2001; 158:1140–1142Crossref, Medline, Google Scholar
83 Levitt JJ, McCarley RW, Nestor PG, et al: Quantitative volumetric MRI study of the cerebellum and vermis in schizophrenia: clinical and cognitive correlates. Am J Psychiatry 1999; 156:1105–1107Medline, Google Scholar
84 Deicken RF, Feiwell R, Schuff N, et al: Evidence for altered cerebellar vermis neuronal integrity in schizophrenia. Psychiatry Res 2001; 107:125–134Crossref, Medline, Google Scholar
85 Andreasen NC, O'Leary DS, Flaum M, et al: Hypofrontality in schizophrenia: distributed dysfunctional circuits in neuroleptic-naive patients. Lancet 1997; 349:1730–1734Crossref, Medline, Google Scholar
86 Crespo-Facorro B, Paradiso S, Andreasen NC, et al: Recalling word lists reveals “cognitive dysmetria” in schizophrenia: a positron emission tomography study. Am J Psychiatry 1999; 156: 386–392Google Scholar
87 Williams RS, Hauser SL, Purpura DP, et al: Autism and mental retardation. Arch Neurol 1980; 37:749–753Crossref, Medline, Google Scholar
88 Bauman M, Kemper TL: Histoanatomic observations of the brain in early infantile autism. Neurology 1985; 35:866–874Crossref, Medline, Google Scholar
89 Ritvo ER, Freeman BJ, Scheibel AB, et al: Lower Purkinje cell counts in the cerebella of four autistic subjects: initial findings of the UCLA-NSAC autopsy research report. Am J Psychiatry 1986; 143:862–866Crossref, Medline, Google Scholar
90 Kemper TL, Bauman M: Neuropathology of infantile autism. J Neuropathol Exp Neurol 1998; 57:645–652Crossref, Medline, Google Scholar
91 Courchesne E, Yeung-Courchesne R, Press GA, et al: Hypoplasia of cerebellar vermal lobules VI and VII in autism. N Engl J Med 1988; 318:1349–1354Crossref, Medline, Google Scholar
92 Murakami JW, Courchesne E, Press GA, et al: Reduced cerebellar hemisphere size and its relationship to vermal hypoplasia in autism. Arch Neurol 1989; 46:689–694Crossref, Medline, Google Scholar
93 Rapin I: Autism in search of a home in the brain. Neurology 1999; 52:902–904Crossref, Medline, Google Scholar
94 Leroi I, O'Hearn E, Marsh L, et al: Psychopathology in patients with degenerative cerebellar diseases: a comparison to Huntington's disease. Am J Psychiatry 2002; 159:1306–1314Crossref, Medline, Google Scholar
95 DelBello MP, Strakowski SM, Zimmerman ME, et al: MRI analysis of the cerebellum in bipolar disorder: a pilot study. Neuropsychopharmacology 1999; 21:63–68Crossref, Medline, Google Scholar
96 Parvizi J, Anderson SW, Martin CO, et al: Pathological laughter and crying: a link to the cerebellum. Brain 2001; 124:1708–1719Crossref, Medline, Google Scholar
97 Annoni JM, Ptak R, Caldara-Schnetzer AS, et al: Decoupling of autonomic and cognitive emotional reactions after cerebellar stroke. Ann Neurol 2003; 53:654–658Crossref, Medline, Google Scholar
98 Bechara A, Damasio H, Tranel D, et al: Deciding advantageously before knowing the advantageous strategy. Science 1997; 275:1293–1295Google Scholar
99 Desmond JE, Fiez JA: Neuroimaging studies of the cerebellum: language, learning and memory. Trends Cogn Sci 1998; 2:355–362Crossref, Medline, Google Scholar
100 Schmahmann JD (ed): The Cerebellum and Cognition. Int Rev Neurobiol 1997; 41Google Scholar
101 Schmahmann JD: Cerebellum and brainstem, in Brain Mapping: The Systems. Edited by Toga A, Mazziotta J. San Diego, Academic Press, 2000, pp 207–259Google Scholar
102 Voogd J, Glickstein M: The anatomy of the cerebellum. Trends Neurosci 1998; 21:370–375Crossref, Medline, Google Scholar
103 Haines DE, Dietrichs E: An HRP study of hypothalamo-cerebellar and cerebello-hypothalamic connections in squirrel monkey (Saimiri sciureus). J Comp Neurol 1984; 229:559–575Crossref, Medline, Google Scholar
104 Andrezik JA, Dormer KJ, Foreman RD, et al: Fastigial nucleus projections to the brain stem in beagles: pathways for autonomic regulation. Neuroscience 1984; 11:497–507Crossref, Medline, Google Scholar
105 Heath RG: Fastigial nucleus connections to the septal region in monkey and cat: a demonstration with evoked potentials of a bilateral pathway. Biol Psychiatry 1973; 6:193–196Medline, Google Scholar
106 Schmahmann JD, Pandya DN: Anatomic organization of the basilar pontine projections from prefrontal cortices in rhesus monkey. J Neurosci 1997; 17:438–458Crossref, Medline, Google Scholar
107 Schmahmann JD, Pandya DN: The cerebrocerebellar system. Int Rev Neurobiol 1997; 41:31–60Crossref, Medline, Google Scholar
108 Schmahmann JD, Pandya DN: Projections to the basis pontis from the superior temporal sulcus and superior temporal region in the rhesus monkey. J Comp Neurol 1991; 308:224–248Crossref, Medline, Google Scholar
109 Schmahmann JD, Pandya DN: Course of the fiber pathways to pons from parasensory association areas in the Rhesus monkey. J Comp Neurol 1992; 326:159–179Crossref, Medline, Google Scholar
110 Schmahmann JD, Pandya DN: Prelunate, occipitotemporal, and parahippocampal projections to the basis pontis in rhesus monkey. J Comp Neurol 1993; 337:94–112Crossref, Medline, Google Scholar
111 Schmahmann JD: From movement to thought: anatomic substrates of the cerebellar contribution to cognitive processing. Hum Brain Mapp 1996; 4:174–198Crossref, Medline, Google Scholar
112 Schmahmann JD: The role of the cerebellum in affect and psychosis. J Neurolinguistics 2000; 13:189–214Crossref, Google Scholar

